

Sinice
Co jsou to ty sinice zač?
Sinice jsou jednobuněčné organismy, které jsou schopny utvářet vláknité stélky. Patří mezi bakterie, prokaryota – to znamená, že v buňce mají jeden chromozom, který není zvlášť obalen dalšími membránami (nemají tedy jádro). Evolučně se pohybujeme u velmi starobylé skupiny – z doložených nálezů víme, že sinice žily již v období před 3,5 – 2,5 miliardy let (prekambrium). Dokonce jim vděčíme za atmosféru obohacenou o kyslík, již během dávné historie postupně přetvářely svojí fotosyntetickou aktivitou. Postupný úbytek živin na rané Zemi znamenal, že organismy byly nuceny hledat alternativní zdroj uhlíku pro výrobu cukrů. A jakmile vývoj přinesl reakční centra citlivá ke sluneční energii, začali předci autotrofních buněk vychytávat jako potenciální zdroj uhlíku oxid uhličitý z prostředí atmosféry. Oproti jiným bakteriím sinice později jako první ovládly mašinérii, která pracuje s elektrony ze štěpené vody. Ostatní bakterie schopné fotosyntézy se totiž naučily oxidovat pouze sirovodík jako zdroj elektronů potřebných k redukci oxidu uhličitého.

Ekologie a význam
Sinice můžeme najít takřka na kterémkoli stanovišti. Osidlují vody sladké, slané, termální či vlhkou půdu stejně jako skalní povrchy, kosterní pozůstatky a kmeny stromů. Výjimku nepředstavují ani jeskyně a jiná temná místa, kde sinice z fotosyntetizujících organismů jasně dominují.
Sinice (i jiné bakterie) a řasy jsou obvykle prvními kolonizátory nově vzniklých substrátů. Jako první zde také utváří organickou hmotu a formují lokální podmínky. Významně se podílejí na tvorbě hornin – podporují jejich zvětrávání produkcí povlaků mnoha polymerů, které se chovají jako vododržný tmel, nebo sami povrch naleptávají. Jako vazači plynného dusíku našly své uplatnění v zemědělství, kde jejich činnost blaze přispívá produktivitě rýžových polí. Poslední dobou začínají být sinice stále více populární jakožto zdroj alternativních bílkovin a jiných živin. V neposlední řadě si s nimi spojujeme problematické vodní květy, které často negativně ovlivňují zisk pitné vody. Jejich krátký život totiž vede k hromadění a následnému rozkladu odumřelé masy u dna, čímž se ochuzují spodní části nádrží o mimořádné množství kyslíku. Svojí působností dokáží zapříčinit rozkolísání režimu pH, se kterým souvisí nárůst škodlivého volného amoniaku – k neblahému vlivu sinic rovněž přispívá vylučování toxických látek.
Někteří zástupci (běžně Nostoc a Anabaena) jsou významnými mutualisty s drobnými houbami, se kterými utvářejí lišejníky (např. Peltigera). Nezřídka také kooperují s některými mechorosty, kapradinami i dokonce nahosemennými rostlinami. U typových symbióz vystupují sinice obvykle jako prostředníci dusíku, který získávají z okolního vzduchu (viz dále). Jako partneři nevstupují přímo do buněk, naopak obsazují prostor mezi nimi (v dutinkách vystlaných slizem), anebo obrůstají houbová vlákna (hyfy).

Morfologie a anatomie
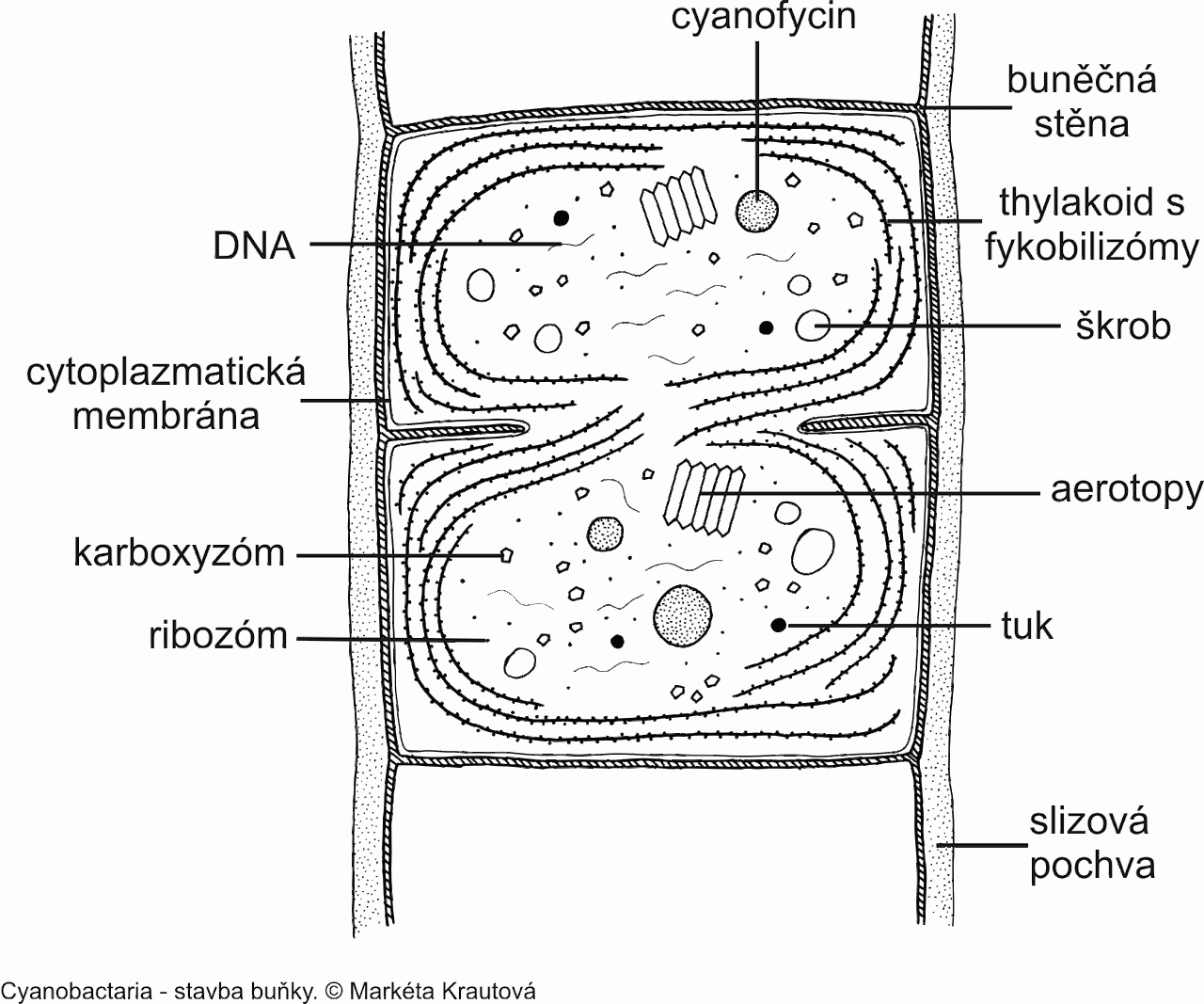
Je třeba si uvědomit, že sinice na rozdíl od řas postrádají zelený plastid – ten vznikl až v momentě, kdy určitá eukaryotická buňka pohltila sinici a nestrávila ji. To, že nedošlo k digesci, bylo evolučně nesmírně důležité, proto existují takové rostliny, které dobře známe. Protoplast sinic obklopuje cytoplazmatická membrána, jejímž vychlípením se zformovaly thylakoidy jako hlavní jednotky fotosyntetického aparátu. Na površích těchto zelených elektráren jsou rozesety bílkovinné fykobilozómy – molekulární antény přispívající sběrem sluneční energie zelenému barvivu chlorofylu.

(Morelli et al. 2021)
Buněčnou stěnu tvoří peptidoglykan (cukr s navázanými aminokyselinami) a několik dalších polysacharidů. Celé buňky k tomu bývají uloženy ve slizové pochvě, která obsahuje mimo mukopolysacharidy navíc pektinové kyseliny, která na sebe váží vodu. Nezřídka jsou součástí gelové vrstvy uhličitanové či železité konkrece, které nosné vnější lešení ještě více utvrzují. Slizové pouzdro i proto nabývá různých barev (od žlutohnědé po fialovou). Protoplazma (neboli tekutý obsah buňky) není vcelku kapalná, z 50 % sušiny ji tvoří bílkoviny. Uvnitř volně plave chromozom (nositel genomu) a někdy rovněž aerotopy, což jsou struktury napomáhající regulovat vztlakový potenciál. Buněčnou výplň dotváří dále kapénky lipidů, sinicový zásobní škrob (alfa-1,4-polyglukan), ribozómy, cyanofycinová zrna (obsahující aminokyseliny arginin a asparagin jako zásobní látky dusíku) a v neposlední řadě karboxyzómy – jsou to anology pyrenoidů řas, čili drobné ústrojky vyplněné klíčovým enzymem RuBisCO, který fixuje CO2 (v malé míře i O2) do energetických molekul. V nadbytku dostupného fosforu jej buňka uskladňuje ve formě tzv. volutinových zrn. I přestože sinice nedisponují bičíkem, jak tomu u bakterií bývá, někteří zástupci se aktivně pohybovat umí (mj. čeleď Oscillatoriaceae) – kloužou se po slizu a odstrkují (ev. přitahují) specifickými kontraktilními vlákny pod membránou.
Sinice nezřídka své buňky modifikují do podoby heterocytů, které bývají větší vůči ostatním buňkám a jsou od nich odděleny póry. Ty se vyznačují velmi mohutnou stěnou, nepropustnou pro jakékoli plyny vyjma dusíku. Zprostředkovávají totiž příjem N2. Plynný dusík si zabudovávají do jiných sloučenin (redukcí do NH3) s pomocí enzymu nitrogenázy . Druhy nevlastnící heterocyty jsou závislé na bytí ve striktně bezkyslíkatém prostředí, protože tento enzym nevystojí již stopy kyslíku (schopnost vázat dusík tak u těchto zástupců spíše zaostává. Kdežto heterocyt propouštějící pouze dusík se jeví jako nejefektivnější volba. Limitující pro všechny vazače bývá dostatečný příjem železa a molybdenu, které jsou v nitrogenáze obsaženy a hrají ústřední roli při fixaci.
Chromatická adaptace – jsou opravdu modrozelené?
Sinice jsou označovány jako modrozelené řasy, ovšem jejich barva doopravdy mate, neboť bývá značně proměnlivá. Celý problém s barevnými tóny často spočívá u již zmiňovaných světlosběrných výběžků, fykobilozomů. Ty jsou složené z pigmentů strukturně příbuzných chlorofylu – obvykle červeného fykoerythrinu a modrého fykocyaninu.
K takzvané chromatické adaptaci přistupují sinice za konkrétních podmínek výživy, například za nedostatku dusíku v prostředí (vzduchu) rozkládají některé vlastní pigmenty, čímž postupně mění svoji barvu. Častější vysvětlení tkví ale v dostupnosti a kvalitě osvětlení. Za zhoršených světelných podmínek umějí buňky měnit rozložení jednotlivých fykobilinů k obrazu svému, aby se absorbční spektra těchto molekul maximálně kryla s přicházejícími paprsky. Všude totiž neproniká stejné bílé světlo, kupříkladu vodní společenstva nemají k dispozici celé spektrum slunečního záření, poněvadž průchod fotonů hustým vodním prostředím vyžaduje více energie. Hladina ještě nemalou část světla rozptyluje (Černý & Petrusek 2022a). Jakmile nevysvětlují konečné zbarvení fykobiliny, pak podněcují pigmentaci asi slizové obaly, do nichž jsou ukládány nejrůznější látky, které hrají mnohdy všemi barvami. Sinice tak můžeme nalézt v odstínech od purpurové až po černou.

(upraveno podle Meene et al. 2006; Dominguez-Martin et. al 2022); autor schématu: © AAT Bioquest
Rozmnožování
Množí se dělením buněk popřípadě celých kolonií. Dělí se od obvodu směrem dovnitř, kdy se u sinic nevláknitých vchlipují spolu s plazmatickou membránou i vrstvy buněčné stěny. Vláknité typy sinic takto využívají jen vnitřní vrstvy stěny, díky čemuž zůstává pro trichom (vlákno) svrchní vrstva společná, aby utvářela souvislý okraj vlákna pro kolonii. Vlákná dokáží produkovat hormogonie, což jsou několikabuněčné podlouhlé útvary, které se oddělují od vlákna mateřského a jsou schopny se smykovaně pohybovat. Za zhoršených podmínek (potravní deprivace, resp. energetický status) formují akinety sloužící právě k přežití nepříznivého období.
Toxiny škodné a neškodné
Do prostředí vylučují množství nejrůznějších látek, ať už odpadních či cíleně podmíněných. Toxiny sinic jsou nerůznější povahy. Apel k vyluzování těchto látek pramení z konkurence, kdy právě takové sloučeniny dokáží potlačit růst jiných. Tyto jedy narušují protivníkovi oxygenní fotosyntézu nebo syntézu RNA. Podle orgánových soustav, které poškozují, se roztřiďují mezi hepatotoxické (poškozující jaterní buňky), neurotoxické (ovlivňující tok neurotransmiterů přímým navázáním na jejich receptor, či třeba blokací sodných kanálů), cytotoxické (bránící syntéze cytochromů a jiných proteinů) nebo toxiny dermatotoxické (poškozující a dráždící pokožku). K dnešku známe nespočet molekulových variant a výzkum struktur těchto jedů i detekce nových toxinů nadále zůstává aktuální. v problematice vodohospodářství.
Zdroje:
van de Meene, A., Hohmann-Marriott, M., Vermaas, W., Roberson, R. (2006): The three-dimensional structure of the Cyanobacterium Synechocystis sp. PCC 6803. Archives of microbiology 184: 259–270. Dostupné z: doi: 10.1007/s00203-005-0027-y
Dominguez-Martin, M., Sauer, P., Kirst, H., Sutter, M., Bína, D., Greber, B., Nogales, E., Polívka, T., Kerfeld, Ch. (2022): Structures of a phycobilisome in light-harvesting and photoprotected states. Nature 609: Dostupné z: doi: 1-11. 10.1038/s41586-022-05156-4